Phytophthora blight, caused by the oomycete pathogen Phytophthora capsici, is one of the most serious diseases threatening pepper production, often leading to widespread plant death and drastic yield losses. However, the key transcription factors regulating resistance to this disease in pepper and their underlying mechanisms remain unclear. Recently, a research team led by Professor Wei Cheng at the College of Life Sciences, Anhui Normal University, published a paper in Biology titled “Three NAC Transcription Factors, CaNAC61, CaNAC79, and CaNAC92, Negatively Regulate Pepper Resistance Against Phytophthora capsici.” Through transcriptomic screening and functional validation, the study identified three NAC transcription factors—CaNAC61, CaNAC79, and CaNAC92—as negative regulators of resistance to Phytophthora blight in pepper, thereby providing new gene targets for disease-resistance breeding.

Based on prior RNA-seq data regarding the interaction between chili pepper and Phytophthora capsici, the research team identified eight differentially expressed NAC genes and focused on CaNAC61, CaNAC79, and CaNAC92. RT-qPCR validation revealed that CaNAC61 and CaNAC79 exhibited early, transient induction—peaking rapidly at 3 hours post-inoculation (hpi) before declining—whereas CaNAC92 showed sustained late-stage induction, becoming significantly upregulated after 24 hpi and maintaining high expression levels through 72 hpi. These distinct temporal dynamics suggest that the three genes may function at different stages of the defense response. Phylogenetic analysis indicated that CaNAC61, CaNAC79, and CaNAC92 cluster into different NAC subfamilies. Multiple sequence alignment confirmed that all three possess a highly conserved N-terminal NAC domain (subdivided into five subdomains, A–E) but exhibit high variability in their C-terminal regions. Subcellular localization assays demonstrated that GFP fusion proteins of all three genes localized specifically to the nucleus, consistent with the typical characteristics of transcription factors. Silencing CaNAC61, CaNAC79, or CaNAC92 individually via virus-induced gene silencing (VIGS) significantly enhanced the chili pepper plants’ resistance to P. capsici; this was evidenced by reduced lesion diameters on detached leaves and a lower disease index following root inoculation. Conversely, transient overexpression of any of these three genes in chili pepper leaves led to significantly larger lesions and markedly suppressed transcript levels of defense marker genes, including CaPR1 (SA pathway) and CaDEF1 and CaLOX1 (JA pathway). Stable overexpression of these genes in Nicotiana benthamiana yielded similar results, with transgenic plants exhibiting hypersusceptibility to the pathogen. Collectively, these findings demonstrate that CaNAC61, CaNAC79, and CaNAC92 act as negative regulators of Phytophthora resistance in chili pepper. The study suggests that these three NAC transcription factors may compromise plant immunity by inhibiting salicylic acid (SA)- and jasmonic acid (JA)-mediated defense signaling pathways. Notably, although all three factors repress CaPR1, CaDEF1, and CaLOX1, they exhibit differential effects on CaPR2 and CaPR10—with CaNAC61 specifically repressing CaPR2 and CaNAC92 specifically repressing CaPR10—suggesting that their functions are not entirely redundant.
This study expands the “negative regulatory module” within the transcriptional regulatory network governing Phytophthora resistance in pepper. The presence of negative regulators is physiologically significant for preventing immune overactivation and maintaining normal growth and development. In breeding applications, the precise downregulation or editing of these negative regulatory genes holds promise as an effective strategy for enhancing resistance to Phytophthora blight in pepper, offering new insights for the development of stable and durable disease-resistant varieties.
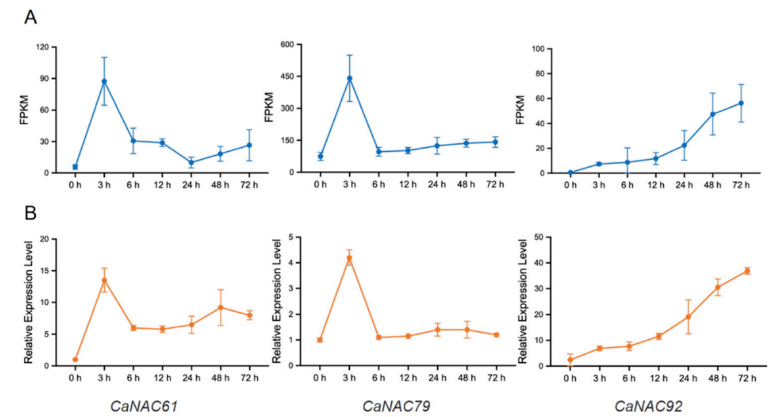
Figure 1. Expression patterns of CaNAC61, CaNAC79, and CaNAC92 during P. capsici infection in pepper: (A) Transcriptional patterns of CaNAC61, CaNAC79, and CaNAC92 at 0, 3, 6, 12, 24, 48, and 72 h post-inoculation with P. capsici were determined by RNA-seq. (B) The expression patterns of CaNAC61, CaNAC79, and CaNAC92 at different time points post-inoculation were validated by RT-qPCR. Data are presented as the mean ± standard deviation (SD) from three independent biological replicates.
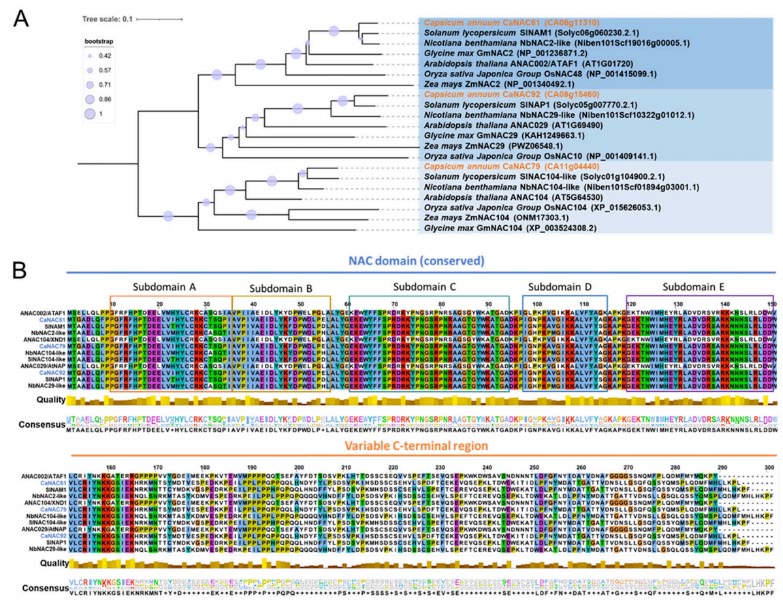
Figure 2. Phylogenetic relationships and conserved domain analysis of CaNAC61, CaNAC79, and CaNAC92: (A) A phylogenetic tree was constructed using NAC protein sequences from Capsicum annuum, Solanum lycopersicum, Nicotiana benthamiana, Glycine max, Arabidopsis thaliana, Oryza sativa, and Zea mays. CaNAC61, CaNAC79, and CaNAC92, which are highlighted in orange, were clustered into distinct phylogenetic clades with representative homologous NAC proteins from other plant species. Bootstrap values are represented by the size of the circles, and the scale bar indicates evolutionary distance. (B) Multiple sequence alignment was performed for CaNAC61, CaNAC79, and CaNAC92 with their homologous proteins. Sequence alignment revealed that all three proteins contain a conserved N-terminal NAC domain, a characteristic feature of the NAC transcription factor family that can be divided into five subdomains.
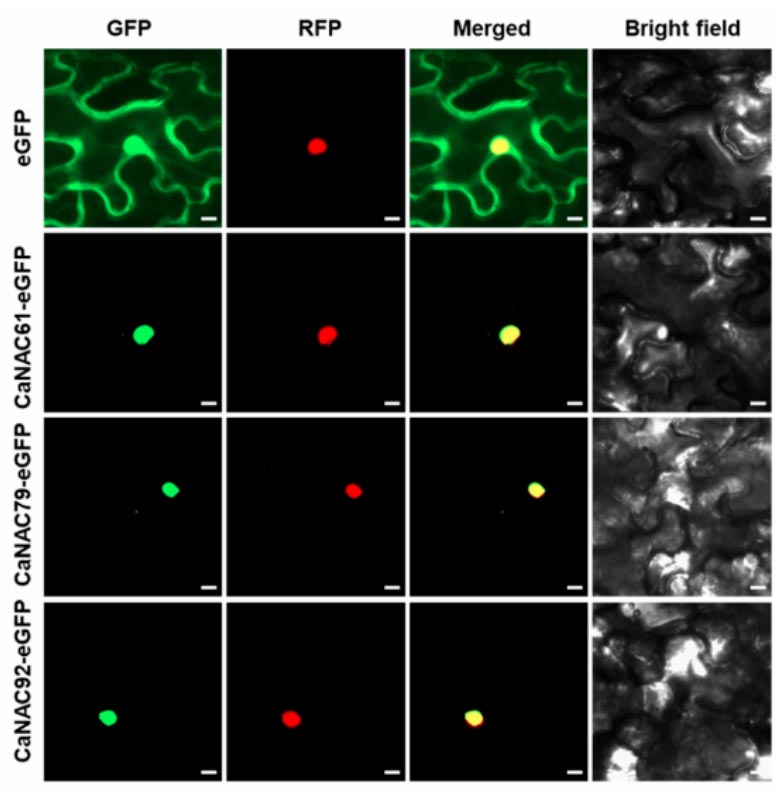
Figure 3. Subcellular localization of CaNAC61, CaNAC79, and CaNAC92. CaNAC61-eGFP, CaNAC79-eGFP, and CaNAC92-eGFP fusion proteins were transiently expressed in N. benthamiana leaves and co-expressed with the nuclear marker H2B-RFP. Confocal microscopy revealed that the GFP signals of CaNAC61, CaNAC79, and CaNAC92 strongly co-localized with the RFP signals, indicating that all three proteins are predominantly localized in the nucleus. eGFP was used as the empty vector control, showing fluorescence signals in both the nucleus and cytoplasm. GFP, green fluorescent protein. RFP, red fluorescent protein. Bar = 10 μm.
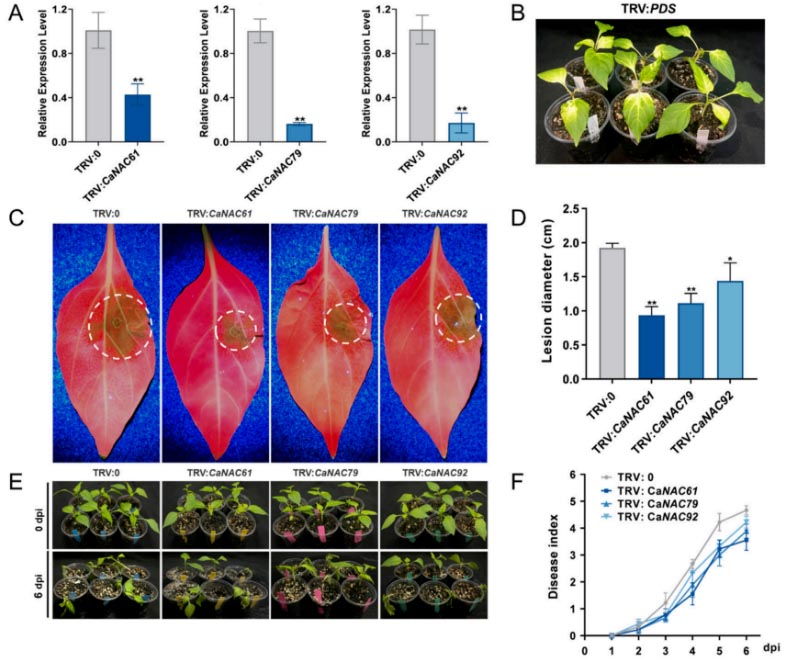
Figure 4. Silencing of CaNAC61, CaNAC79, and CaNAC92 enhances resistance to P. capsici in pepper: (A) Transcript levels of CaNAC61, CaNAC79, and CaNAC92 in TRV-mediated gene-silenced pepper plants were measured by RT-qPCR, with TRV: 0 serving as the negative control. Data represent the mean ± SD of three independent biological replicates. (B) Approximately 3–4 weeks post-agroinfiltration, TRV: PDS-treated plants exhibited typical photobleaching, confirming the effectiveness of the VIGS system. (C) Representative disease symptoms of detached VIGS leaves at 3 days post-inoculation (dpi) with P. capsici zoospores. (D) Lesion diameters after P. capsici inoculation at 3 dpi. (E) Representative disease symptoms of TRV-treated pepper plants subjected to root inoculation at 0 dpi and 6 dpi. (F) Disease indexes of these TRV-treated pepper plants after root inoculation with P. capsici zoospores. Data are presented as the mean ± SD from six independent biological replicates. Asterisks indicate significant differences based on Student’s t-test (* p < 0.05, ** p < 0.01).
Figure 5. Transient overexpression of CaNAC61, CaNAC79, and CaNAC92 enhances susceptibility of pepper leaves to P. capsici infection: (A) Transcript levels of CaNAC61, CaNAC79, and CaNAC92 in transiently overexpressing pepper leaves were measured by RT-qPCR. The empty vector (EV) served as the control. Data represent the mean ± SD of three independent biological replicates. (B) Lesion diameters in pepper leaves at 2 days post-inoculation (dpi) with P. capsici zoospores. Data are presented as the mean ± SD from six independent biological replicates. (C) Representative disease symptoms of pepper leaves following P. capsici inoculation at 2 dpi. (D) Expression of defense marker genes in pepper leaves transiently overexpressing CaNAC61, CaNAC79, or CaNAC92. Transcript levels of CaPR1, CaDEF1, and CaLOX1 were determined by RT-qPCR at 2 days post-agroinfiltration. Data were obtained from three independent biological replicates. The empty vector (EV) served as the control. All data are presented as the mean ± SD, and asterisks indicate significant differences based on Student’s t-test (* p < 0.05, ** p < 0.01).
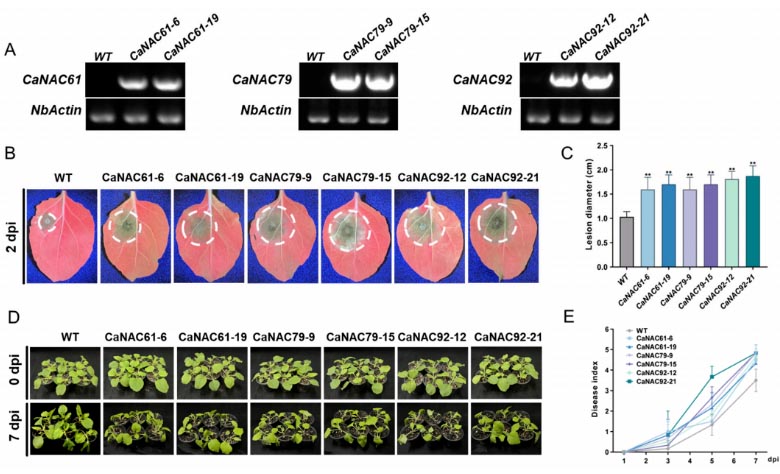
Figure 6. Ectopic stable overexpression of CaNAC61, CaNAC79, and CaNAC92 enhances susceptibility to P. capsici in N. benthamiana: (A) RT-PCR analysis of CaNAC61, CaNAC79, and CaNAC92 transcript levels in transgenic N. benthamiana lines overexpressing each of these genes. NbActin was used as the internal reference, and wild-type (WT) plants served as the control.. (B) Representative disease symptoms of detached leaves from different transgenic N. benthamiana lines following P. capsici inoculation at 2 dpi. (C) Lesion diameters of these transgenic N. benthamiana leaves after P. capsici inoculation at 2 dpi. (D) Representative disease symptoms of these transgenic N. benthamiana plants subjected to root inoculation at 0 dpi and 7 dpi. (E) Disease indexes of different transgenic N. benthamiana lines after root inoculation with P. capsici zoospores. The data are presented as the mean ± SD from six independent biological replicates. Asterisks indicate significant differences based on Student’s t-test (** p < 0.01).
Disclaimer: The information is sourced from industry research on chili peppers; please contact us for removal in the event of copyright infringement.