Bacterial wilt, a devastating soil-borne disease caused by Ralstonia solanacearum, seriously threatens pepper production. Plants need to activate an immune response to defend against pathogen infection, but over-activation can inhibit growth; therefore, precise regulation of immune strength is crucial. Recently, a research team from Fujian Agriculture and Forestry University and other institutions revealed the molecular mechanism by which the C2H2 type zinc finger protein CaZAT11 negatively regulates pepper immunity against bacterial wilt by interacting with the HD-Zip transcription factor CaHDZ27, and competitively binds to CaHDZ27 with the 14-3-3 protein CaTFT7, thereby achieving a dynamic balance between growth and defense. The related research findings, titled “CaZAT11 Negatively Fine-Tunes Pepper Immunity to Ralstonia solanacearum by Repressing CaHDZ27,” were published in the academic journal Plant, Cell & Environment.

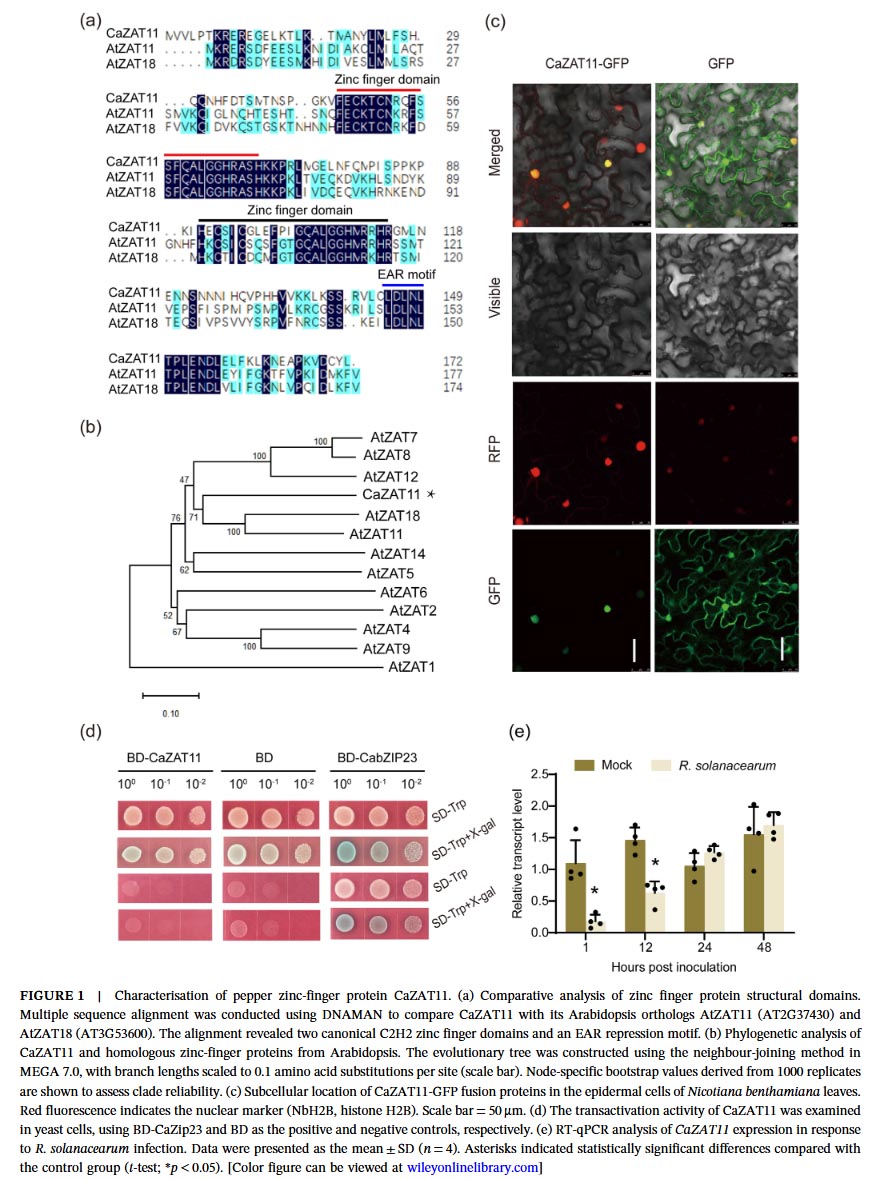
This study first screened for the candidate protein CaZAT11, which interacts with the positive immune regulator CaHDZ27 in pepper, using GST pull-down combined with mass spectrometry analysis. Sequence analysis showed that CaZAT11 contains two typical C2H2 zinc finger domains and a C-terminal EAR repressive motif, classifying it as a C2H2-type zinc finger transcription factor. Subcellular localization indicated that CaZAT11 is located in the cell nucleus, and yeast self-activation assays confirmed its lack of transcriptional activation activity, consistent with the characteristics of a transcriptional repressor. Expression analysis showed that CaZAT11 expression was downregulated in the early stages of Ralstonia solanacearum infection (1-12 hours), and in different resistant materials, the expression level of CaZAT11 in susceptible materials was higher than that in resistant materials, suggesting its potential negative regulation of immunity.
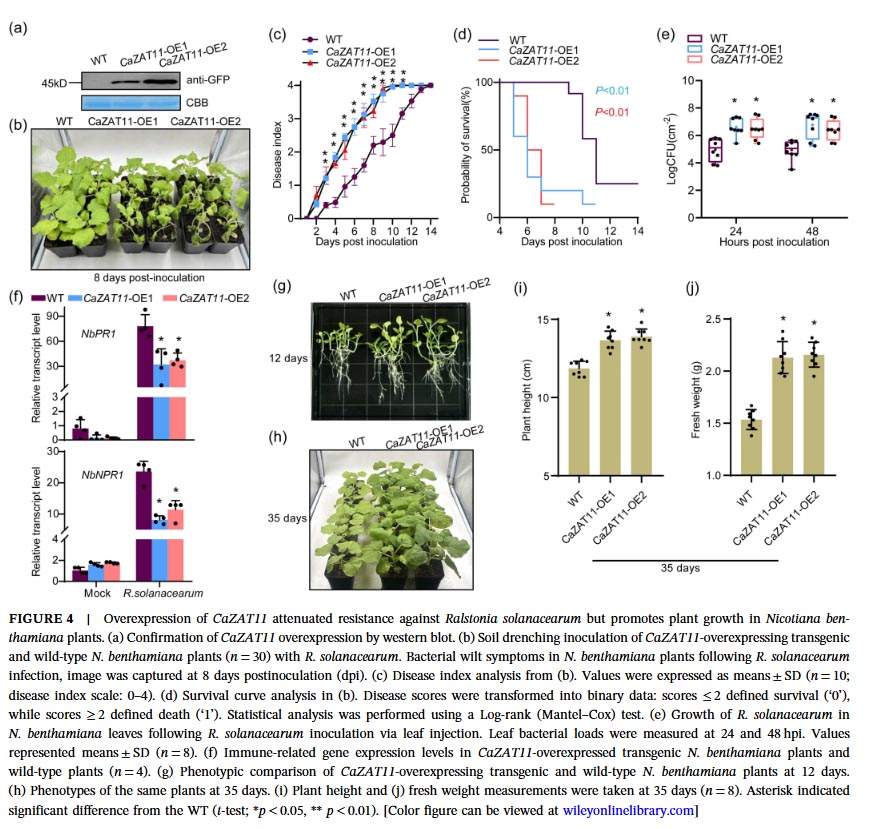
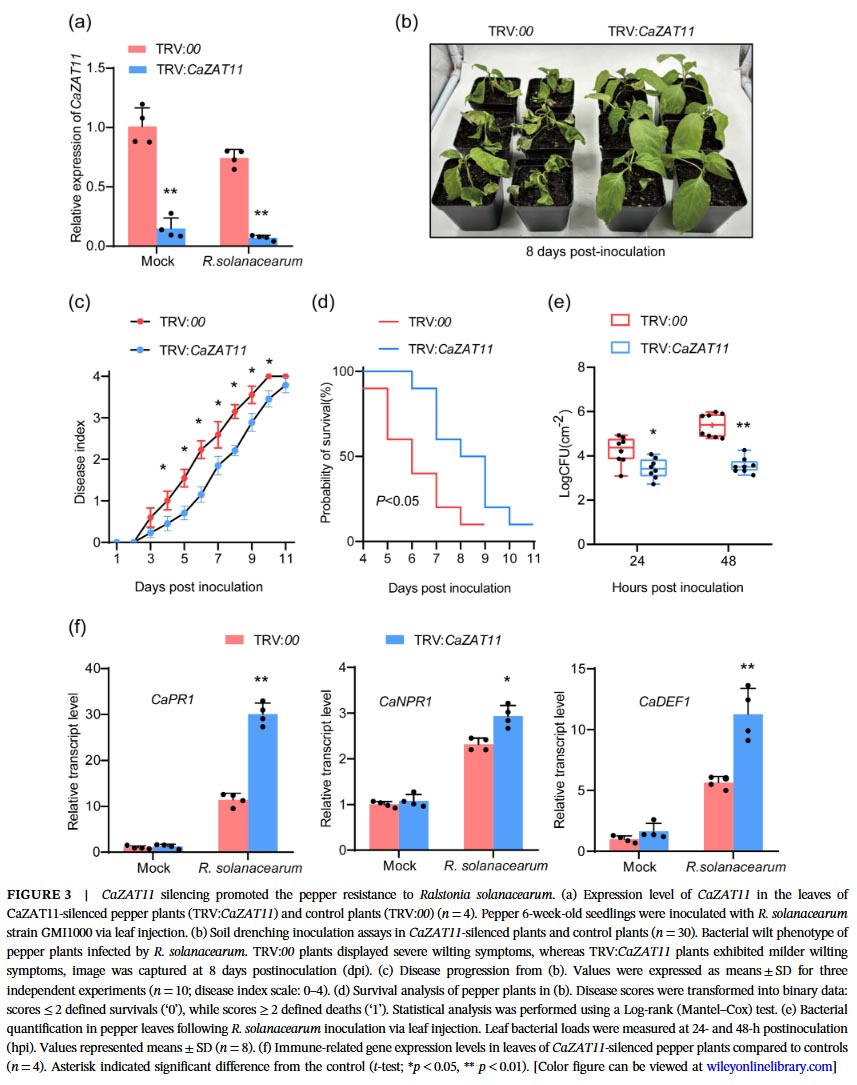
Silencing CaZAT11 using virus-induced gene silencing technology significantly enhanced the resistance of pepper plants to bacterial wilt, reduced the disease index and mortality rate, suppressed bacterial proliferation, upregulated the expression of immune-related genes (CaPR1, CaNPR1, CaDEF1), and did not affect plant growth. Conversely, overexpression of CaZAT11 in tobacco increased plant susceptibility to bacterial wilt, accelerated bacterial proliferation, and downregulated expression of immune genes, but simultaneously promoted plant growth (increased plant height and fresh weight), indicating that CaZAT11 promotes growth while suppressing immunity.
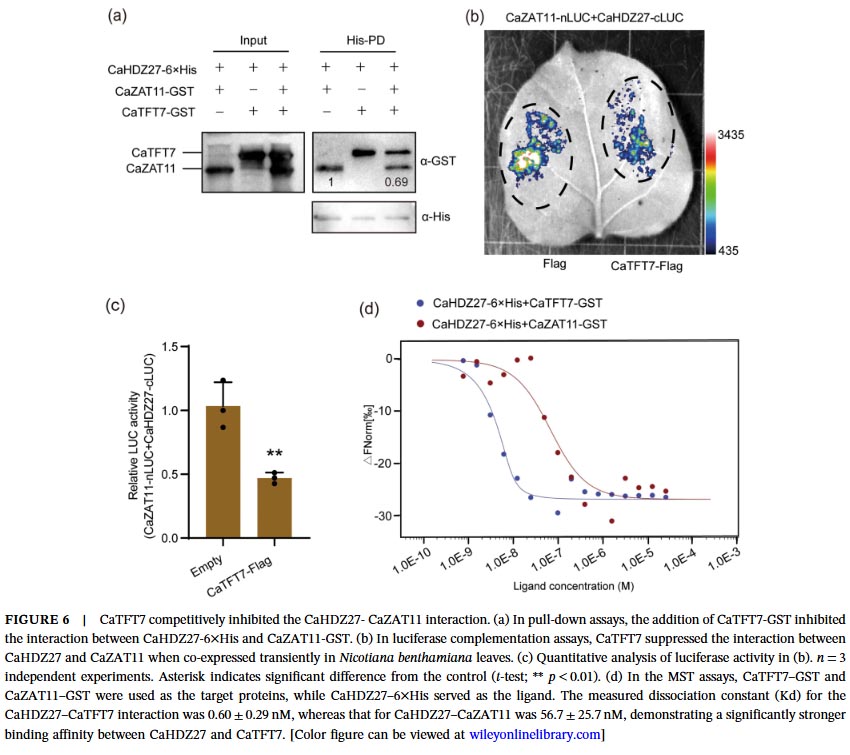
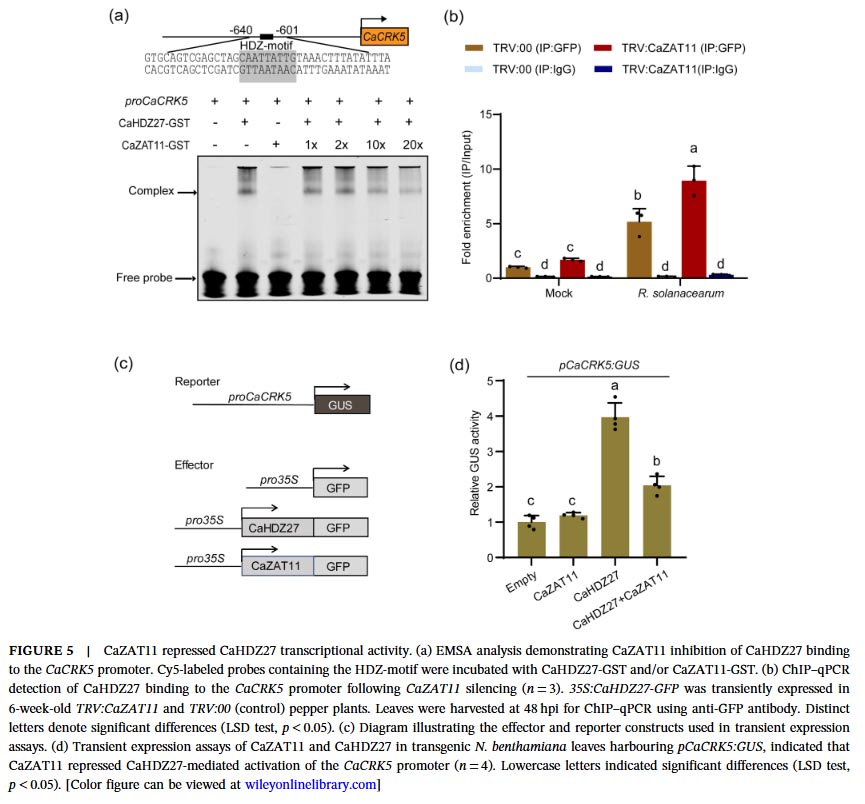
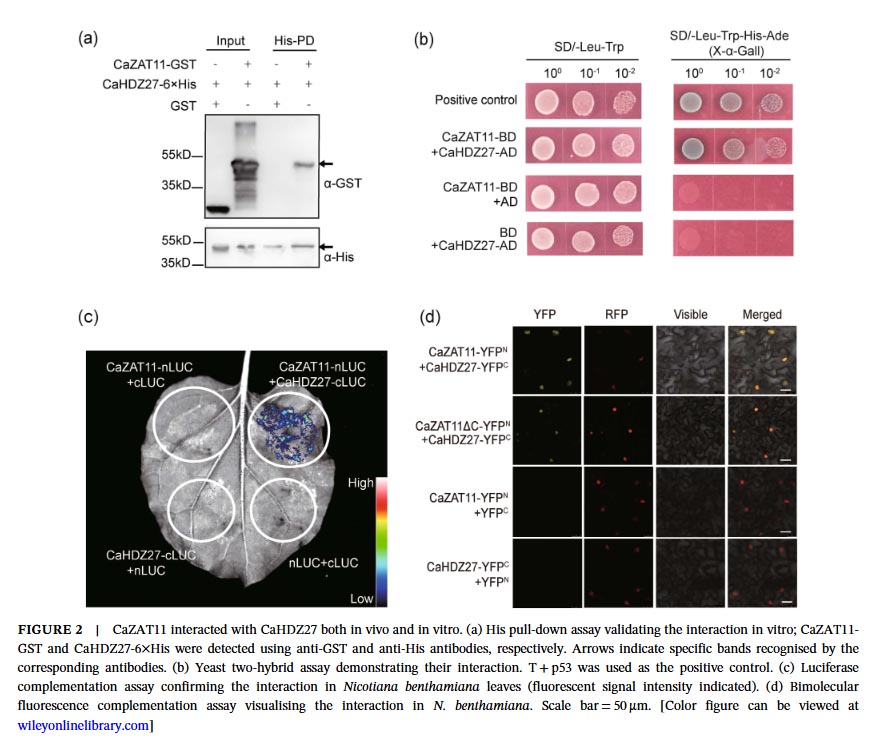
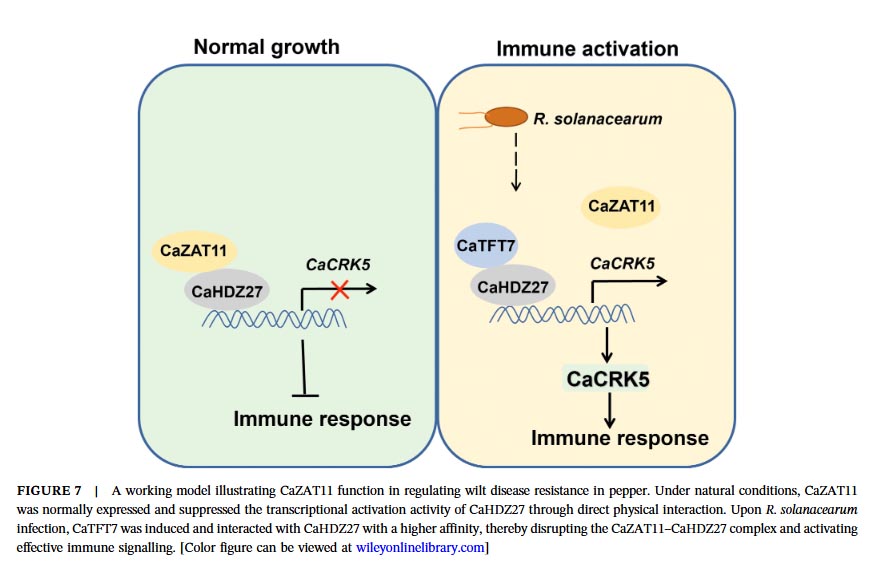
Molecular mechanism studies revealed physical interactions between CaZAT11 and CaHDZ27 both in vitro and in vivo. Under normal conditions, CaZAT11 inhibits the transcriptional activation activity of CaHDZ27 through its EAR motif, weakening the binding of CaHDZ27 to its target gene CaCRK5 promoter, thereby inhibiting CaCRK5 expression and maintaining immune silencing. Gel migration assays and chromatin immunoprecipitation-quantitative PCR confirmed that CaZAT11 protein dose-dependently inhibits the binding of CaHDZ27 to the CaCRK5 promoter, and silencing CaZAT11 enhances this binding. Notably, the CaZAT11ΔC variant lacking the EAR motif loses its ability to inhibit CaHDZ27 transcriptional activity, and overexpression of this truncated form does not alter plant susceptibility, indicating that the EAR motif is essential for CaZAT11’s inhibitory function.
Further research revealed that the previously reported immunomodulator CaTFT7 (14-3-3 protein) has a significantly higher affinity for CaHDZ27 than CaZAT11 (Kd value: 0.60 nM vs 56.7 nM). Following Ralstonia solanacearum infection, CaTFT7 is induced to express and preferentially binds to CaHDZ27, competitively dissociating the CaZAT11-CaHDZ27 complex. Simultaneously, CaZAT11 protein levels decrease, thereby relieving inhibition and activating the CaHDZ27-mediated immune response.
In summary, this study proposes a dynamic regulatory model: under normal conditions, CaZAT11 binds to CaHDZ27, inhibiting its transcriptional activity, preventing excessive immune activation, and allocating resources to growth; after pathogen infection, CaTFT7 is induced to bind to CaHDZ27 with higher affinity, replacing CaZAT11, releasing the activity of CaHDZ27, activating downstream immune genes (such as CaCRK5), and initiating disease resistance. This study reveals the crucial role of CaZAT11 as a “brake” molecule in finely regulating the trade-off between immunity and growth, providing new genetic resources and theoretical basis for balancing growth and defense in crop disease resistance breeding.